By: Patricia cartwright
Writing about the quirks of human nature.
Overview: The social brain hypothesis (SBH), a theory developed by Robin Dunbar, attempts to answer the age-old question of why the human neocortex has evolved as large as it has in comparison to other species such as fish, birds, reptiles, and other large mammals (Dunbar & Shultz, 2007). The central argument of this hypothesis lies in the idea that the evolution of the human brain can be largely attributed to social cognitive factors (Dunbar et al. 2007; Shultz & Dunbar, 2007). In other words, the human brain has transformed into a social brain (Adolphs, 2009). The SBH further posits that due to the size of social groups and the complexities that lie in one’s social environment such as social bonding, reciprocal altruism, coalition frequency, deception, perspective taking, mating strategies, and social learning the neocortex in humans has been pushed to evolve (Dunbar & Shultz, 2007; van Schaik et al., 2012). Participation in social groups are understood to be demanding and complex due to the intricacies involved. The maintenance of group cohesion and behaviors that fit in with social group norms become increasingly difficult as the social group size increases (Shultz & Dunbar, 2007). Therefore, to be adequately able to thrive within growing social groups, selection warranted brain evolution and improvements in human social intelligence.
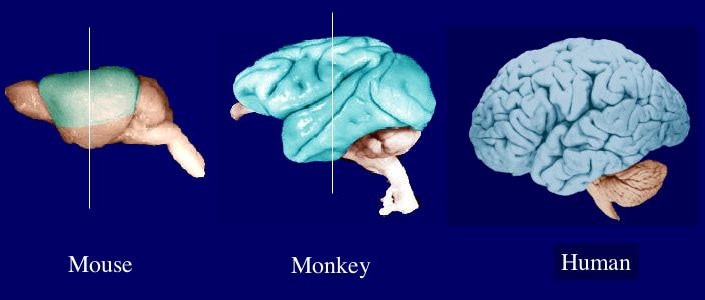
If it’s bigger, it’s better.
Evidence supporting SBH:
Despite the evidence refuting the SBH, it prevails as a widely-cited explanation for brain evolution for several reasons. First, the hypothesis has received increasing empirical support that has found the size of the neocortex to be strongly associated with indices of social complexities (Dunbar & Shultz, 2007). For instance, the relationship cited by Dunbar between neocortex size and group size of various mammals has been replicated by other researchers in the field (Finlay & Darlington, 1995; Byrne & Corp, 2004). Secondly, given the expensive energy costs associated with utilizing higher cognitive brain functions, it is a well-reasoned argument that there is some beneficial social advantage that has driven brain evolution (Dunbar et al., 2007).
Evidence supporting SBH:
Despite the evidence refuting the SBH, it prevails as a widely-cited explanation for brain evolution for several reasons. First, the hypothesis has received increasing empirical support that has found the size of the neocortex to be strongly associated with indices of social complexities (Dunbar & Shultz, 2007). For instance, the relationship cited by Dunbar between neocortex size and group size of various mammals has been replicated by other researchers in the field (Finlay & Darlington, 1995; Byrne & Corp, 2004). Secondly, given the expensive energy costs associated with utilizing higher cognitive brain functions, it is a well-reasoned argument that there is some beneficial social advantage that has driven brain evolution (Dunbar et al., 2007).
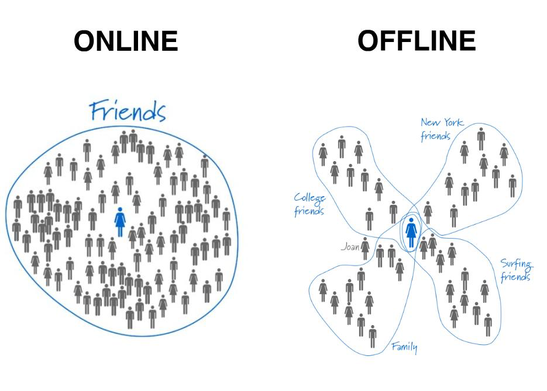
Dunbar’s number theory seems to hold even in the face of the commonalities like cellphones and social networks such as Facebook and LinkedIn (Kanai et al. 2012; MacCarron et al., 2016). Although the introduction of social networks should have debunked Dunbar’s number, they have instead strengthened the theory. Researchers have found that the size and complexity of online social networks are significantly positively correlated with and reflected in grey matter density in focal regions in the cortex (i.e., left MTG, right STS, & right entorhinal cortex) and amygdala (Bickart et al., 2010; Kanai et al., 2012). Moreover, despite the influence of social networks and the presence of outliers in each direction, the figure of 150 prevails as far as representing the limit of genuine social relationships one can have (MacCarron et al., 2016).
Critiques and counterpoints: In extant literature, the SBH is widely favored as an explanation for the evolution of large brains in humans and primates, over traditional explanations based on ecological or developmental problem solving (Dunbar & Shultz, 2007). The traditional explanations of the evolution of the human brain had a tendency to narrowly emphasize the need for specific abilities such sensory and technical competencies (e.g., way-finding or mental mapping and scavenging) as the reason behind the human brain expansion (Dunbar et al. 2007). Although the SBH is more comprehensive in nature than traditional explanations, it contains several shortcomings that should be brought to light.
The SBH, for instance, cannot adequately account for grade shifts or why certain species such as Orangutans and aye ayes possess unusually large brains. This is particularly surprising given the simplistic structure and nature of their social hierarchy and environments (van Schaik et al. 2012). Moreover, the SBH does not provide an explanation for why mammalian lineages have significantly smaller brains but possess equal socio-cognitive abilities and function in equally complex relationship dynamic societies (van Schaik et al. 2012). Likewise, when looking at other species such as small-brained fish, the SBH is not able to account for the ability of this species to exhibit socio-cognitive abilities that match that of primates (van Schaik et al. 2012). Moreover, in Tim Ingold’s critique of the SBH, he points out that the data supporting the SBH has been selective and confirmatory in nature. Although the SBH may not be able to adequately explain brain size for all mammalian lineages, it may be the best explanation we have for humans and primates.
The Magic Number
One of the two final counterpoints to be considered is the argument that the complexities of social relationships may be due to the evolution of mammalian intelligence, rather than the demands an increase in social group size brings. Intelligent individuals bring their own challenges, so it is reasonable to argue that this relationship may be flipped. A second and final counterpoint to be considered is with Dunbar’s number. This theory posits that 150 is the cognitive limit of people that one person can maintain stable relationships with. Since Dunbar’s research on this theory, Dunbar’s number has been found to have an upper limit, which does not necessarily invalidate Dunbar’s number but it calls it into question (Kanai et al. 2012; McCarty et al. 2001; MacCarron et al., 2016).
Critiques and counterpoints: In extant literature, the SBH is widely favored as an explanation for the evolution of large brains in humans and primates, over traditional explanations based on ecological or developmental problem solving (Dunbar & Shultz, 2007). The traditional explanations of the evolution of the human brain had a tendency to narrowly emphasize the need for specific abilities such sensory and technical competencies (e.g., way-finding or mental mapping and scavenging) as the reason behind the human brain expansion (Dunbar et al. 2007). Although the SBH is more comprehensive in nature than traditional explanations, it contains several shortcomings that should be brought to light.
The SBH, for instance, cannot adequately account for grade shifts or why certain species such as Orangutans and aye ayes possess unusually large brains. This is particularly surprising given the simplistic structure and nature of their social hierarchy and environments (van Schaik et al. 2012). Moreover, the SBH does not provide an explanation for why mammalian lineages have significantly smaller brains but possess equal socio-cognitive abilities and function in equally complex relationship dynamic societies (van Schaik et al. 2012). Likewise, when looking at other species such as small-brained fish, the SBH is not able to account for the ability of this species to exhibit socio-cognitive abilities that match that of primates (van Schaik et al. 2012). Moreover, in Tim Ingold’s critique of the SBH, he points out that the data supporting the SBH has been selective and confirmatory in nature. Although the SBH may not be able to adequately explain brain size for all mammalian lineages, it may be the best explanation we have for humans and primates.
The Magic Number
One of the two final counterpoints to be considered is the argument that the complexities of social relationships may be due to the evolution of mammalian intelligence, rather than the demands an increase in social group size brings. Intelligent individuals bring their own challenges, so it is reasonable to argue that this relationship may be flipped. A second and final counterpoint to be considered is with Dunbar’s number. This theory posits that 150 is the cognitive limit of people that one person can maintain stable relationships with. Since Dunbar’s research on this theory, Dunbar’s number has been found to have an upper limit, which does not necessarily invalidate Dunbar’s number but it calls it into question (Kanai et al. 2012; McCarty et al. 2001; MacCarron et al., 2016).
Most theories tend to test one research question before ascertaining whether alternative, equally plausible explanations could also make the same predictions (Dunbar & Shultz, 2007). |
Conclusion: I would argue that the SBH, despite its shortcomings, is the best explanation we have available for the evolution of the neocortex among humans and primates. Although the SBH is unable to account for key components such as grade shifts, the evidence supporting the SBH is widely cited, such as studies showing the positive correlation between human neocortex volume and grey matter density as well as the aversive effects of social deprivation on child brain development and on prisoners (Kanai et al., 2012). Given this evidence, its popularity among anthropologists and other scientists is warranted but perhaps it is time to consider a broader hypothesis that can account for the aforementioned shortcomings of the SBH in addition to providing an explanation for the evolution of large brains in all mammalian lineages and other species.
References/Additional Resources:
- Adolphs, R. (2009). The social brain: Neural basis of social knowledge. Annual Review of Psychology, 60, 693–716. http://doi.org/10.1146/annurev.psych.60.110707.163514
- Bickart, K. C., Wright, C. I., Dautoff, R. J., Dickerson, B. C., & Barrett, L. F. (2011). Amygdala volume and social network size in humans. Nature Neuroscience, 14, 163-164. doi:10.1038/nn.2724
- Byrne, B., & Corp, N. (2004). Neocortex size predicts deception rate in primates. Biological Sciences, 271, 1693-1699.
- Dunbar, R. I. & Shultz, S. (2007). Evolution in the social brain. Science, 317, 1344–1347.
- Finlay, B. L. & Darlington, R. B. (1995). Linked regularities in the development and evolution of mammalian brains. Science, 268, 1578-1584.
- Kanai, R., Bahrami, B., Roylance, R., & Reese, G. (2012). Online social network size is reflected in human brain structure. Biological Sciences, 279, 1327-1334. doi:10.1098/rspb.2011.1959
- Lihoreau, M., Latty, T., & Chittka, L. (2012). An exploration of the social brain hypothesis in insects. Frontiers in Physiology, 3, 1-7. http://doi.org/10.3389/fphys.2012.00442
- MacCarron, P., Kaski, K., & Dunbar, R. (2016). Calling Dunbar's numbers. Social Networks, 47, 151-155. doi:10.1016/j.socnet.2016.06.003
- McCarty, C., Killworth, P. D., Bernard, R., Johnsen, E. C., and Shelley, G. A. (2001). Comparing two methods for estimating network size. Human Organization, 1, 28-39.
- Shultz, S., & Dunbar, R. I. (2007). The evolution of the social brain: Anthropoid primates contrast with other vertebrates. Biological Sciences, 274, 2429–2436. http://doi.org/10.1098/rspb.2007.0693
- van Schaik, C. P., Isler, K., & Burkart, J. M. (2012). Review: Explaining brain size variation: from social to cultural brain. Trends In Cognitive Sciences, 16, 277-284. doi:10.1016/j.tics.2012.04.004
- van Vugt, M. (2012). The missing link: Leadership, identity, and the social brain. British Journal Of Psychology, 103, 177-179.






 RSS Feed
RSS Feed

{kind=link}